
Modelling the spatial structure of chromatin
The focus of our group is on the understanding of the spatial structure of the genome, its regulation and its impact on genomic activity.
In the nucleus of higher order cells DNA is complexed with proteins forming so called nucleosomes, cylindrical units, that are connected by DNA forming a bead-on-a-string like structure. This structure folds into chromatin. The spatial structure of chromatin can mediate contacts between or occlude access to the genomic regions. We developed a coarse-grained computer model of chromatin that can be used to model chromatin structures up to the size of several mega base pairs and the formation of topologically associating domains. Currently we collaborate in the DFG priority program SPP2202.
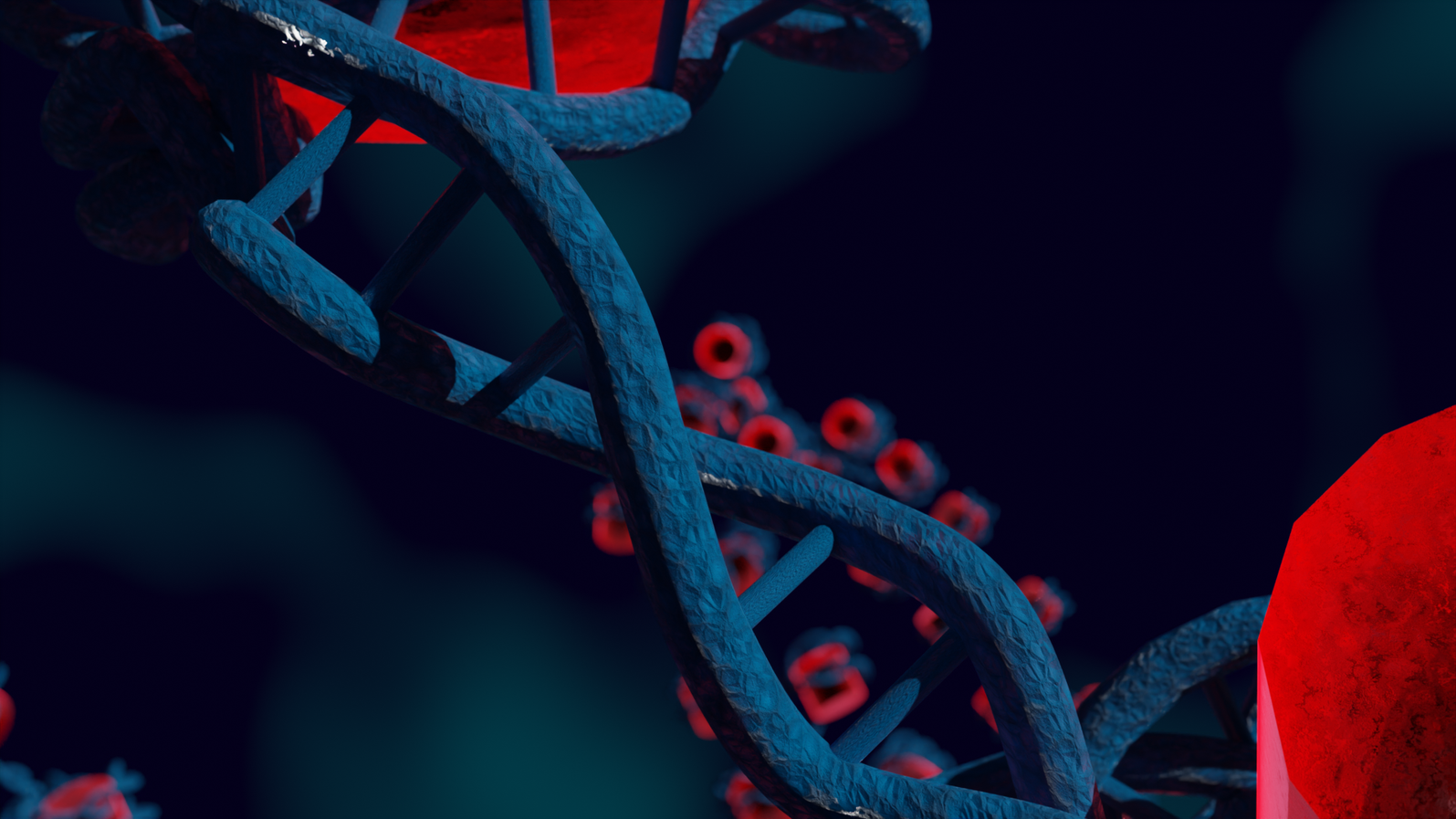
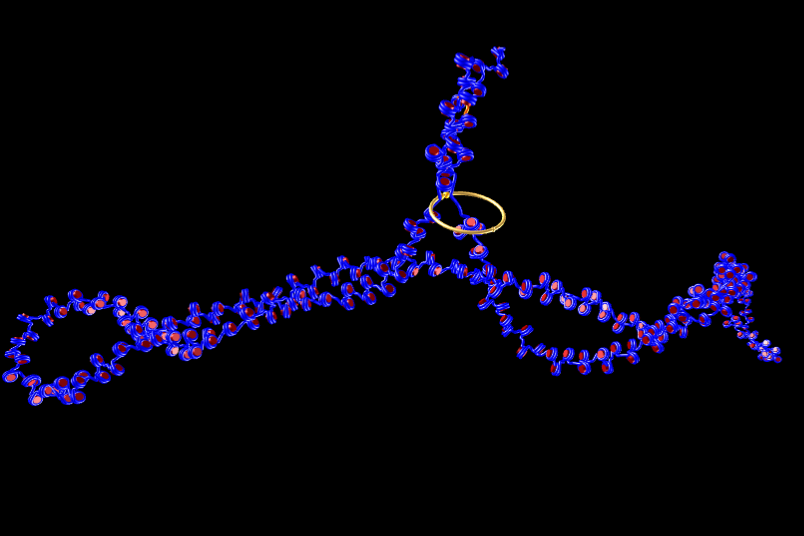
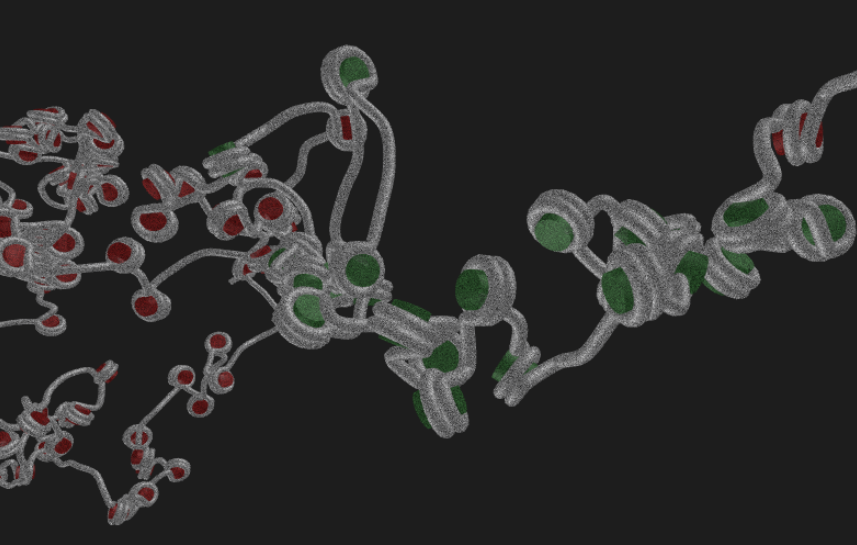
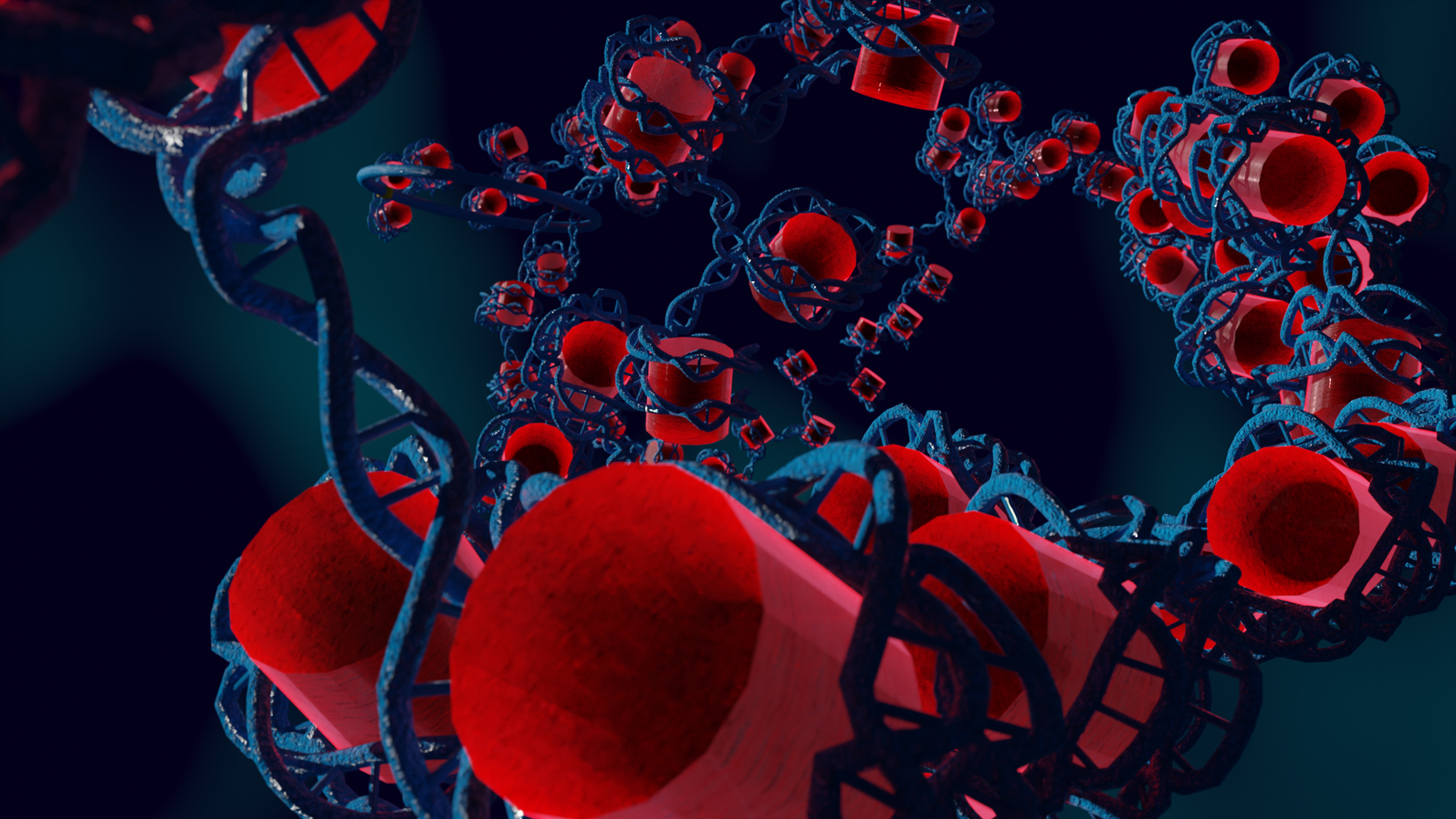
Highlights of our research
- The model can reproduce the experimental properties of reconstituted chromatin and chromatin from chicken erythrocytes. (Stehr, 2008) (Stehr, 2010) The model can explain force extension curves of reconstituted chromatin (Kepper, 2011)
- It is important to utilize replica exchange not plain Metropolis Monte Carlo to avoid sampling artifacts. (Stehr, 2008)
- The strength of the nucleosome-nucleosome interaction directs fiber compaction, fiber conformation, and higher order folding. (Stehr, 2008)
- Variations of the lengths of linker DNA induce large changes of the fiber conformation. (Kepper, 2008)
- Even the relocation of a single nucleosome has strong changes on the geometry of regular spaced polynucleosomes. (Müller, 2014)
- In transcriptional active and inactive regions the number of nucleosomes, the amount of bound H1 and the strength of internucleosomal interaction varies strongly. (Brandstetter, 2021)
Modelling Metastatic Progression
Cancer is one of the most common causes of death. Mostly, metastasis is leading to death. We developed a computer model that describes the spread of malign cells in various organs and different interventions. This software is Open Source available. This work was done in close collaboration with Prof. Dr. Udo Schumacher, University Medical Center Hamburg-Eppendorf.
Highlights of our research
- Our model can reproduce data from a patient with a hepatocellular carcinoma. (Bethge, 2012)
- Resection, chemotherapy and radiation therapy can be modelled quantitatively (Bethge, 2015)
- In xenograft models it is important to determine the number of surviving engrafted cells to properly analyze the data. Assuming the same number for all tumors result in large errors. (Hoffmann, 2020)
- Natural killer cells induce dormancy of metastasis (Brodbeck, 2014). The molecular mechanism was experimentally clarified by another group. (Lopes, 2021)
- Chemotherapy changes the vessel tree geometry in the primary tumor, most notably the fractal dimension. This has impact on the spreading of malign cells. (Frenzel, 2017)
Funding
